- Гематологические исследования
- Биохимические исследования
- Биохимическое исследование крови и мочи
- Специфические белки в сыворотке крови и моче
- Биохимические исследования пункционной жидкости
- Биохимические исследования кала
- Биохимические исследования наследственных нарушений обмена веществ
- Исследования при мочекаменной болезни
- Витамины, микроэлементы, оксидативный стресс
- Жирные кислоты
- Фракция трансферрина при алкоголизме
- Неинвазивные маркеры заболеваний печени
- Химический анализ камней
- Эндокринологические маркеры
- Гормоны, участвующие в углеводном обмене
- Гормоны, участвующие в процессе роста
- Гормоны, секретируемые адипоцитами
- Маркеры фертильности
- Маркеры нормальной / патологической беременности
- Гормоны надпочечников
- Гормоны щитовидной железы
- Нейрогормоны
- Пренатальный скрининг на аномалии плода
- Ренин-ангиотензин-альдостероновая система
- Онкологические маркеры
- Маркеры вирусных инфекций
- Маркеры сердечно-сосудистых патологий
- Исследование анемий
- Маркеры патологии костной системы
- Маркеры аутоиммунных болезней
- Антиспермальные антитела
- Аутоантитела при эндокринных, сердечных, почечных заболеваниях
- Аутоантитела при неврологических заболеваниях
- Аутоантитела при дерматологических заболеваниях
- Аутоантитела при пернициозной анемии
- Аутоантитела при сахарном диабете
- Маркеры аутоиммунных заболеваний печени и желудочно-кишечного тракта
- Маркеры ревматических заболеваний и васкулитов
- Маркеры для наблюдения за развитием и лечением болезней
- Маркеры антифосфолипидного синдрома
- Серологические исследования инфекционных болезней
- Аллергологические и иммунологические исследования
- Молекулярно-биологические исследования
- Цитогенетические исследования
- Микробиологические исследования
- Токсикология
- Цервико-вагинальная цитология
- Гистопатологические исследования
- Генетическое консультирование
- Uncategorized


Общая информация
Генетика основного комплекса гистосовместимости
Основной комплекс гистосовместимости (MHC) представляет собой генную группу, расположенную на короткой руке хромосомы 6, которая производит поверхностные клеточные маркеры, которые играют важную роль в процессе распознавания: молекулы класса I и класса II. У людей молекулы MHC класса I включают лейкоцитарные антигены (HLA = антиген лейкоцитов человека) HLA-A, HLA-B и HLA-C, тогда как молекулы MHC класса II включают HLA-DR, HLA-DQ и HLA-DP. Эти молекулы составляют классические антигены, участвующие в трансплантации. На уровне MHC также кодируются молекулы класса III: MHC-связанные компоненты комплемента (C2, C4 и Bf), 21-гидроксилаза (CYP21), высокочувствительный белок (Hsp) 70 и фактор некроза опухоли (TNF).
MHC растягивается в длину 3600 кб и включает в себя 224 идентифицированных гена, из которых прогнозируется экспрессия в количестве 128; около 40% экспрессируемых генов имеют функцию, связанную с иммунной системой. Гены HLA расположены в 6 суб-областях, которые следуют друг за другом в следующем порядке: HLA-A, HLA-C, HLA-B, HLA-DR, HLA-DQ, HLA-DP, HLA-A дистально от центромеры. Каждая субрегиона кодирует по крайней мере один поверхностный гликопротеин (см. Рисунок 1).
Рисунок 1: последовательность генов HLA I и II на хромосоме 6
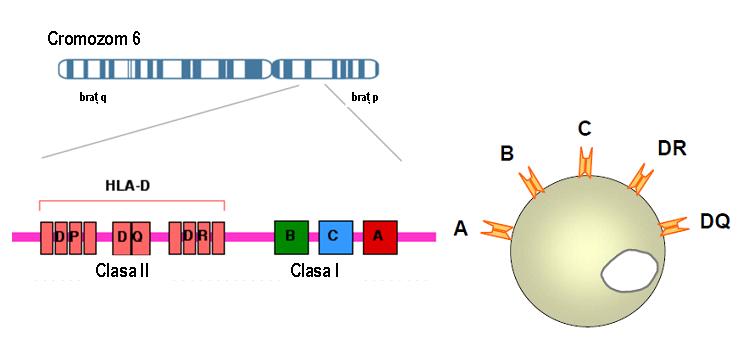
За одним исключением, гены HLA характеризуются высоким полиморфизмом, точнее, каждый ген показывает несколько аллелей в популяции. Учитывая важную роль системы HLA в иммунном ответе, представляется, что этот полиморфизм необходим для выживания видов и сохраняется в популяции путем отбора.
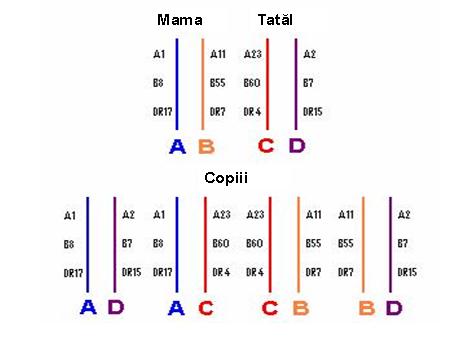
Гены MHC тесно связаны друг с другом, что означает массовую сегрегацию в ходе мейоза. Комплекс связанных генов, которые расположены на одной из пар гомологичных хромосом и которые массово сегрегируют у потомства, называется гаплотипом. Каждый человек наследует два гаплотипа HLA-по одному от каждого родителя-и, таким образом, имеет два аллеля для каждого гена, экспрессирующего кодоминант (см. Рисунки 2,3).
Рисунок 2: аллели-унаследованные пары от мамы и папы для каждого локуса HLA
Наблюдение, что аллели в различных генетических локусах встречаются в популяции в пределах одного и того же гаплотипа, причем частота значительно выше, чем предполагалось только на случайной основе, определяет дисбаланс связей – особенность системы HLA – которая простирается от HLA-A до HLA-DQ включительно. Наиболее известным дисбалансом связи является гаплотип A1, Cw7, B8, DR17(3), DR52, DQ2, встречающийся в европеоидной популяции с частотой, в 4 раза превышающей ожидаемую 2.
Рисунок 3: Как передаются гаплотипы HLA детям семьи
Роль молекул HLA класс I и II
Классические молекулы, относящиеся к классу I MHC, называемые у людей HLA-A, HLA-B и HLA-C, представляют собой гетеродимеры, состоящие из гликозилированного трансмембранного полипептида, связанного с β2-микроглобулином, который экспрессируется на всех ядросодержащих клетках и владеет ключевые роли в иммунном ответе:
● Показывает пептидные антигены, полученные из цитоплазматических белков ( например, вирус или аномальный белок в злокачественной клетке) клональных рецепторов, специфичных для каждого пептида CD8+Т-лимфоцитов. Эти пептиды возникают в результате переваривания белков на уровне лизосом, обычно их длина составляет около 9 аминокислот. Каждый аллотип MHC I выбирает свои собственные Т-клетки; таким образом, существует 6 подмножеств Т-клеток, каждая из которых специализируется на аллотипах HLA-A, HLA-B и HLA-C материнского или отцовского происхождения. После распознавания пептидов CD8 + Т-клетки лизируют клетки-мишени.
● Их экспрессия на поверхности клеток выполняет защитную функцию в врожденном иммунном ответе, предотвращая лизацию клеток-мишеней NK-клетками (”естественный убийца"). В частности, NK могут распознавать и лизировать клетки-мишени, которые не экспрессируют молекулы HLA класса I на своей поверхности, тем самым играя важную роль в надзоре против вирусов и опухолевых клеток. Уровень экспрессии молекул HLA класса I на поверхности клеток максимален на лимфоидных клетках; в ходе иммунного ответа экспрессия увеличивается под действием цитокинов (интерферон γ и TNF); с другой стороны, опухоли и некоторые вирусы (например, ВИЧ) могут подавлять экспрессию HLA I.
Наряду с классическими генами в область MHC I также включены гены со сниженным полиморфизмом, обозначенные как HLA-E, HLA-F и HLA-G2;8.
Молекулы, относящиеся к классу II MHC, HLA-DR, HLA-DQ и HLA-DP, представляют собой гетеродимеры, состоящие из двух нековалентно связанных трансмембранных гликопротеинов – α-цепи (33-35 КДА) и β-цепи (26-28 КДА).
Каждая из 3 субрегионов, DR, DQ и DP, включает по крайней мере один ген A (который кодирует α-цепь) и один ген B (который кодирует β-цепь). Субрегион DR кодирует одну или две молекулы DR, в зависимости от гаплотипа. Он содержит один экспрессируемый ген DRA, который похож на несколько гаплотипов. Ген DRB, наиболее близкий к центромеру, DRB1, кодирует высоко полиморфную β-цепь, которая, связанная с α-цепью, образует молекулу класса II, которая преобладает на поверхности клеток и которая экспрессирует серологические специфичности DR1-DR18. Второй экспрессируемый ген DRB, который присутствует только в определенных гаплотипах, локализуется между локусами DRB1 и DRA и, в зависимости от экспрессируемых аллелей DRB1, называется DRB3, DRB4 или DRB5. Полученные молекулы DR имеют серологические особенности DR52, DR53 и DR51 соответственно. Субрегион DQ включает полиморфные гены DQA1 и DQB1, которые кодируют гетеродимер DQ ce, несущий серологические специфичности DQ1-9. Субрегион DP содержит 2 набора генов A и B: набор dpa1 и dpb1 с высоким полиморфизмом кодирует белковый продукт DP; другой набор состоит из псевдогена 2.
В отличие от молекул класса I, молекулы класса II экспрессируются только на поверхности антигенпрезентирующих клеток (APC): моноцитов, макрофагов, дендритных клеток, В-лимфоцитов и, кроме того, на активированных Т-лимфоцитах и эпителиальных клетках
Молекулы класса II показывают CD4 + хелперные Т-лимфоциты пептидные антигенные фрагменты, обработанные из экзогенных антигенов, таких как бактерии. Антигенные рецепторы CD4 + Т-лимфоцитов взаимодействуют с антигенным фрагментом комплекса-молекулами HLA класса II и вызывают активацию клеток. Как и в случае с классом I, распознавание антигенного фрагмента эффекторными Т-клетками зависит от полиморфизма аллелей MHC. Таким образом, эффекторная Т-клетка специфична для антигенного пептида в сочетании с конкретным аллельным продуктом класса II, который первоначально представлял антиген (ограничение MCH). После распознавания антигена каждая активированная CD4+ Т-клетка стимулирует дифференцировку в-клеток в продуцирующие антитела плазмоциты и способствует дифференцировке других Т-лимфоцитов в цитотоксические клетки или с супрессорными функциями.
Рисунок 7: представление антигенных фрагментов Т-клеток молекулами HLA I и II, экспрессируемыми на APC
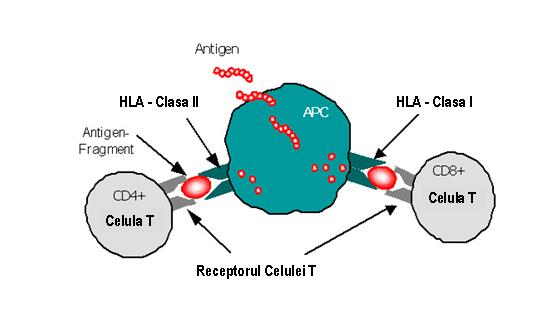
Atat moleculele de clasa I cat si cele de clasa II pot lega alternativ peptide proprii („self”) provenite din degradarea fiziologica a proteinelor celulare. In mod obisnuit acestea nu declanseaza activarea limfocitelor T deoarece sunt tolerate de sistemul imun individual; in unele situatii insa se pot declansa reactii imune care initiaza un proces de distrugere a self-ului ce conduce la autoimunitate2.
Как молекулы класса I, так и молекулы класса II могут поочередно связывать собственные пептиды („self”), возникающие в результате физиологической деградации клеточных белков. Обычно они не вызывают активацию Т-лимфоцитов, потому что они переносятся отдельной иммунной системой; однако в некоторых ситуациях могут быть вызваны иммунные реакции, которые инициируют процесс самоуничтожения, приводящий к аутоиммунитету 2.
Номенклатура HLA
Номенклатура HLA разработана комитетом ВОЗ, созданным для этого. Первоначальная идентификация молекул HLA включала использование аллозеров человека, полученных в результате аллоиммунизации, полученной во время беременности, или выполнения различных трансплантаций. Таким образом, возникающая в результате номенклатура отражает историю этих усилий по открытию, которые устанавливаются в рамках международных семинаров. Аллотипы были обозначены числами, такими как HLA-A2, HLA-B27 и HLA-Cw6, а числа антигенов HLA-A и HLA-B не перекрываются (см. Рисунок 4). Буква "w", первоначально использовавшаяся для обозначения определенной категории семинаров, была сохранена для аллотипов HLA-C с целью отличить их от компонентов комплемента C2 и C4, которые также отображаются в MHC8.
Рисунок 4: Серологический анализ Tipaj HLA
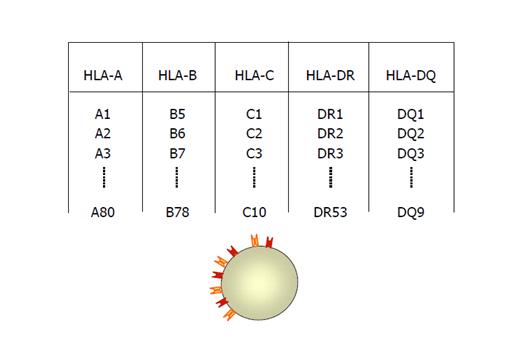
При серологическом тестировании некоторые аллозеры взаимодействуют с более чем одним аллельным продуктом HLA, явление, называемое инкрустированной реактивностью. В случае аллельных продуктов, соответствующих локусам HLA-A и HLA-B, это явление было тщательно изучено и использовано для классификации молекул на антигенные группы, которые реагируют инкрустированно (CREGs). Молекулы класса I в CREG имеют один или несколько антигенных детерминант, которые не восстанавливаются на уровне молекул другого CREG. Антигенный детерминант (эпитоп), который встречается у членов CREG, называется общественной специфичностью (например: HLA-Bw4 и HLA-Bw6)2.
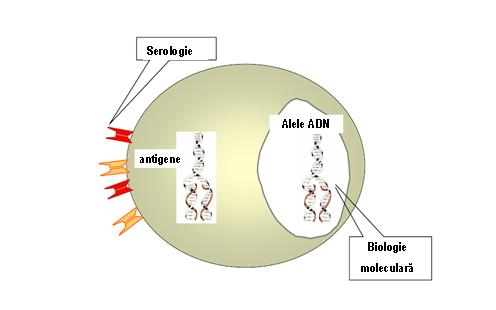
Серологические методы HLA-типа все чаще заменяются методами молекулярной биологии (электрофорез нуклеиновых кислот, методы гибридизации, а также секвенирование), которые привели к изменению номенклатуры с учетом аллелей ДНК. Таким образом, одна серологически определенная специфичность может быть выражена двумя или более (>25) различными аллелями (см. Рисунок 5).
Рисунок 5: типаж HLA-от серологии до молекулярной биологии
В номенклатуре на основе ДНК каждый аллель HLA обозначается названием локуса гена, за которым следует Звездочка * и 4-7-значным числом, указывающим на аллель (например, a*0201 является аллелем гена HLA-A, А B*2701 является аллелем гена HLA-B). Первые 2 цифры в названии каждого аллеля часто основаны на серологическом типе полученной молекулы и/или сходстве нуклеотидных последовательностей с другими аллелями в группе (см. Рисунок 6).
Рисунок 6: тип HLA через молекулярную биологию

Некоторые аллели отличаются последовательностью экзонов ДНК, но не полученной аминокислотной последовательностью (из-за молчаливых или синонимичных замен); эти аллели идентифицируются путем добавления 5-й цифры к 4 общим цифрам (например, B*27051 и B*27052). Кроме того, цифры 5-7, добавленные в обозначение, указывают на то, что 2 аллеля отличаются только последовательностями на уровне интронов (например, DRB4* 0103101 и DRB4* 0103102). В данном примере DRB4* 0103102 показывает сайт сплайсинга модифицированной РНК, который приводит к потере экспрессии аллеля; аллели, которые не экспрессируются как белки, могут иметь N, добавленный к их названию (например, A*215N и DRB4* 0103102n)2;3.
В отличие от серологических методов, HLA-типирование на основе ДНК показывает чистый преимущества:
● он специфичен (каждый реагент для набора четко определен и основан на определенной нуклеотидной последовательности);
● он гибкий (по мере открытия новых аллелей можно получать новые реагенты);
● он более надежен, чем другие методы (не требует жизнеспособных лимфоцитов);
● может широко применяться;
● может различать аллели ДНК, которые специфичны для белков HLA, которые не могут быть дифференцированы серологически; например, человек, представляющий аллель DRB1* 0401, имеет тот же серологический тип DR4, что и человек, который является носителем аллеля DRB4* 04122.
Молекулярные методы идентифицированы в настоящее время 4447 аллелей HLA класса I и II: 965 HLA-A, 1543 HLA-B, 626 HLA-C, 9 HLA-E, 21 HLA-F, 46 HLA-G, 855 HLA-DRB, 35 HLA-DQA1, 107 DQB1, 28 HLA-DPA1, 138 HLA-DPB3. Новые обнаруженные аллели представлены в периодических отчетах комитета ВОЗ, а нуклеотидные последовательности всех аллелей хранятся в компьютерной базе данных (GenBank, EMBL, IMGT/HLA databases)2.
С апреля 2010 года вступает в силу изменение номенклатуры HLA: в результате увеличения количества аллелей HLA было решено ввести": "в название аллелей для разграничения отдельных полей. Таким образом:
A*01010101 → A*01:01:01:01
A*260101 →A*26:01:01
A*3301 → A*33:01
B*0808N → B*08:08N
DRB1*01010101 → DRB1*01:01:01:01
Также ожидаются другие изменения, в том числе то, что буква „w” будет вычтена из названия аллелей HLA-C, но будет сохранена в антигенах HLA-C4.
Заболевания, связанные с системой HLA
Существует значительное количество состояний, которые демонстрируют связь с генами MHC; в большинстве случаев это аутоиммунные заболевания, которые не имеют классического Менделевского способа передачи. Например, анкилопоэтический спондилит связан в 95% случаев с HLA-B27.
Одной из основных проблем, связанных с ассоциациями генов MHC, является их неполная пенетрантность, что делает официальные исследования сегрегации и линкинга очень трудными, если не невозможными, для выполнения. Другим неизвестным является количество аллелей, предрасполагающих к определенному заболеванию в конкретной популяции; кажется, что многие заболевания, связанные с системой HLA, являются полигенными.
Генетическая ассоциация отражена в термине "относительный риск", который представляет собой статистическое соотношение между индивидуальным риском заболевания у носителей генетического маркера и риском у людей тех же групп населения, у которых нет этого маркера. Относительный риск выше 1 указывает на положительную связь между HLA и этим заболеванием 6.
Среди состояний, связанных с HLA, которые были тщательно изучены, есть сахарный диабет 1 типа. Самая сильная связь - это аллели класса II HLA DR и DQ. Гетерозиготный генотип DR3 / DR4 повышает риск развития диабета, за которым следуют гомозиготные генотипы DR4 и DR3.
У европеоидной расы гетеродимеры HLA-DQ (Альфа-цепи DQA1 и бета-цепи DQB1), кодируемые аллелями DQA1*0301, DQB1*0302, DQA1*0501, DQB1*0201, демонстрируют наибольшую связь с диабетом 1 типа. Было отмечено, что DQB1*0302 отличается от DQB1*0301 в положении 57, где отсутствует остаток аспарагиновой кислоты. Такое же отсутствие аспарагиновой кислоты в положении 57 также происходит в аллеле DQB1*0201, и было высказано предположение, что остальная аминокислота будет участвовать в молекулярном механизме восприимчивости к диабету3.
Другие гаплотипы генов HLA класса II играют защитную роль в развитии диабета: DQA1*0102/DQB1*0602/DRB1*1501. В частности, аллель DQB1*0602 обеспечивает защиту даже у родственников пациентов с диабетом первой степени, у которых присутствуют антитела к островковым клеткам поджелудочной железы9.
Абсолютный риск развития диабета 1 типа в зависимости от генотипа HLA показан в таблице ниже 1:
| DR3/DR3 | 1/125 |
| DR3/DRX | 1/500 |
| DR4/DR4 | 1/147 |
| DR4/DRX | 1/476 |
| DR3/DR4 | 1/42 |
| DRX/DRX | 1/5565 |
Тестирование HLA полезно для оценки риска развития диабета у людей с положительным семейным анамнезом.
Пациенты с маркерами HLA dqb1 * 0201 и / или *0302 или предрасполагающими аллелями DR4 (DRB1*0405, DRB1*0402) должны наблюдаться биохимически и иммунологически 5.
В результате внедрения методов на основе ДНК в тип HLA были выявлены некоторые четкие связи между прогрессированием инфекции ВИЧ-1 до синдрома приобретенного иммунодефицита (СПИД) и некоторыми антигенами HLA класса II. Таким образом, в настоящем HLA-B27 наблюдается отсутствие прогрессирования до долгосрочного СПИДа, тогда как связь с HLA-B35 обозначает быстрое прогрессирование. Разница между двумя аллелями заключается в различном количестве вирусных пептидов, которые могут связываться с 2 экспрессируемыми молекулами HLA-B; молекула HLA-B27 связывает 15 различных типов пептидов, происходящих из оболочки вируса, тогда как ни один вирусный пептид не содержит аминокислотных последовательностей, которые предпочтительно связываются с HLA-B35. Следовательно, загрузка молекул HLA пептидами с целью представления антигена представляет собой критическую стадию. ВИЧ-инфекция контролируется CD8+ Т-клетками в течение переменного периода времени; размер этого периода зависит от количества распознанных вирусных пептидов 8.
Что касается связи HLA с боррелиозом, согласно данным литературы сообщается:
– HLA, связанные с рефрактерной болезнью Лайма при лечении, то есть с аутоиммунитетом, вызванным патогеном:
DR1 (HLA-DRB1*0101)
DR2 (HLA-DRB1*1501)
DR4 (HLA-DRB1*0401,*0402,*0403,*0404,*0405,*0407)
– HLA, связанные с низкой выработкой специфических антител Borrelia при наличии определенной инфекции:
alele DR1 (HLA-DRB1*0101,*0102,*0103,*0104,*0105).
В заключение, определение антигенов HLA может быть использовано сегодня в качестве маркера для дифференциальной диагностики многих состояний. В таблице ниже приведены примеры состояний, связанных с определенными созвездиями HLA, наряду с относительным риском заболеваний.
| Заболевание | Антиген HLA | Относительный риск |
| Аллергия на латекс | DR4, DQB1*0302 (DQ3) | 2.4 |
| Аллергия на инсулин | B7, B21 | 5.4 |
| Очаговая алопеция | B12 | 5.4 |
| Экзогенный аллергический альвеолит | DR6 | 16.5 |
| Апластическая анемия | DR2 | 2.2 |
| Пагубная анемия | DR5 | 5.4 |
| Хронический ювенильный артрит | DR8 DR5 | 8.0 3.3 |
| Псориатический артрит | B27 B38 | 10.7 9.1 |
| Реактивный артрит-инфекции: Шигелла, Иерсиния, сальмонелла | B27 | 40.0 |
| Артрит Лайма (инфекция Боррелиоз) | DR2/DR4 | 22.0 |
| Аспириновая астма | DQ2 | 4.1 |
| Болезнь Аддисона (идиопатическая) | DR3 | 6.3 |
| Болезнь Альцгеймера | B7/Cw3 | 28.0 |
| Базедова болезнь | DR3 | 4 |
| Болезнь Бехчета | B51 | 6.3 |
| Болезнь Бехчета( мукокутанный) | B12 | 5.6 |
| Болезнь Бехтерева | B27 | 87.4 |
| Целиакия | DQ2/DQ7/DQ8 | 52.0 |
| Болезнь Крона | DR1, DR4, DR7 | 2.0-7.0 |
| Смешанное заболевание соединительной ткани | DR4 | 3 |
| Первичный билиарный цирроз (ПБЦ) | DR3, DRB1*0803 (DR8) | 3-6.8 |
| Дробьевидная хориоретинопатия или хориоретинопатия Бирдшота | A29 | 224 |
| Первичный склерозирующий холангит (ПСХ) | DR3, DR8 | 4.5-15 |
| Герпетиформный дерматит Дюринга | DR3/DR7 | 17.3 |
| Дерматомиозит | DR3/DR52 | 5 |
| Сахарный диабет 1-го типа | DQ2/DQ3 подтипы DRB1*0405 (DR4)DQ*0602 (DQ6)DQB1*0602 (DQ6) | > 50 Защитный |
| Селективный дефицит иммуноглобулина А (IgА) | DR3 | 17.0 |
| Мембранозный гломерулонефрит | DR3 | 12.0 |
| Гранулематоз Ве́генера | DR9 | 6.7 |
| Идиопатический гемохроматоз | A3/B7/B14 | 90.0 |
| Аутоиммунный хронический гепатит | DR3, DR4, DR8, DR52 | 4.7 |
| Хронический вирусный гепатит В | B35 | 158.0 |
| Здоровые носители» HBsAg | B41 | 11.2 |
| Врожденная гиперплазия надпочечников | B47 DR3 | 15.4 6.3 |
| ВИЧ – Повышенная чувствительность к абакавиру | B * 5701 | 117 |
| вич – медленно прогрессирующее заболевание | B27 B57 | Protector Protector |
| Красный плоский лишай | DR1 | 11.8 |
| Холелитиаз | A19 | 131 |
| Системная красная волчанка (СКВ) | DR2/DR3 | 5.8 |
| Лекарственная волчанка | DR4 | 5.6 |
| Миастения гравис | DR3 DR5 | 3.9 3.6 |
| Нарколепсия | DQB1*0602, DRB1*1501 | 130.0 |
| Волчаночный нефрит | DR2/DQ1 | 14.0 |
| IgA-нефропатия | DR4 | 5.5 |
| Мембранозная нефропатия (МН) | DR3 | 3-12 |
| Подострый склерозирующий панэнцефалит | A29 | 3.6 |
| Пемфигус Вульгарис (обыкновенная пузырчатка) | DR3, DR4 | 14.4 |
| Эритематозная пузырчатка (себорейная пузырчатка, синдром Сенира-Ашера) | DR1 | 7.3 |
| Ревматоидный артрит | DR1/DR4 подтип | 10.2 |
| Ревматическая полимиалгия | DR3 DR4 | 2.3 5.7 |
| Полимиозит | DR3/DR52 | |
| Бляшечный или вульгарный псориаз | B17, B37 Cw6 | 6.7 33.0 |
| Идиопатическая тромбоцитопеническая пурпура | DR2 | 9.2 |
| Синдром Рейтера | B27 | 37.0 |
| Диабетическая ретинопатия | B8 | 4.0 |
| Саркоидоз | B7/B8DR2, DR3, DR5 | 8.5 |
| Саркома Капоши | DR5 | 5.3 |
| Шизофрения | A9/B27 A2/A11 | 11.9 9.8 |
| Склеродермия (PSS) | DR3 DR5 | 16.7 10,0 10.0 |
| Рассеянный склероз | DR2/DQ6 подтипы | 4.1 |
| Андрогенитальный синдром-поздняя форма начала | B14 | 48.5 |
| Андрогенитальный синдром с сольтеряющей формой | B47 | 51.0 |
| Антифосфолипидный синдром | DR4, DR7 | 5.1 |
| Синдром астмы вызванный анальгетиками | A1/B8/DR3 | 28.9 |
| Лимитированная склеродермия(CREST синдром) | DR5 | 8.1 |
| Синдром Гудпасчера | DR2 | 15.9 |
| Синдром Шегрена | DR2/DR3 | 9.7 |
| Рецидивирующий афтозный стоматит | B7 | 6.0 |
| Аутоиммунный тиреоидит(Тиреоидит Хашимото, боезнь Хашимото) | DR3, DR5 | 3.2 |
| Подострый тиреоидит (ПТ) (тиреоидит де Кервена, гигантоклеточный тиреоидит, гранулематозный тиреоидит) | B35 | 13.7 |
| Послеродовой тиреоидит | DR4/DR5 | 8.0 |
| Неонатальная трансиммунная (аутоимунная) тромбоцитопения | DR3 | 9.2 |
| Задний острый увеит | B27 | 10-50 |
Подготовка пациента-специальная подготовка не требуется – в форме отправки будет указано предполагаемое состояние, для которого требуется тестирование HLA7.
Собранный образец-венозная кровь 7.
Контейнер для сбора-vacutainer, содержащий ЭДТА в качестве антикоагулянта7.
Собранное количество-насколько позволяет вакуум7.
Причины отторжения образца-использование гепарина в качестве антикоагулянта; коагулированные или гемолизированные образцы; охлажденные или замороженные образцы7.
Стабильность образца - 7 дней при 2-8ºC7.
Метод – метод-реакция цепной полимеризации (ПЦР) с обнаружением электрофорезом в агарозном геле 7.
Интерпретация результатов
Положительный результат на антиген HLA, связанный с определенным состоянием, показывает генетическую предрасположенность. Это приводит к относительному риску заболеть носителем по сравнению с „нормальным” населением (см. таблицу). В случае подозрения на конкретное состояние выделение антигена HLA является диагностическим критерием7..
Библиография
1. E. Albert Reece, Donal R.Coustan, Steven G.Gabe. Diabetes in Women, ed.3, 2004, 61-62.
2. H. Davis, Richard A. McPherson. Human Leukocyte Antigen: The Major Histocompatibility Complex of Man. In Henry’s Clinical Diagnosis and Management by Laboratory Methods, Saunders-Elsevier, 21st Edition, 8, 876-886.
3. HLA Nomenclature. IMGT/HLA database. www.ebi.ac.uk/imgt/hla/nomenclature/index.html. Ref Type: Internet Communication.
4. HLA Nomenclature Changes Effective April 1, 2010. www.ashi-hla.org. Ref Type: Internet Communication.
5. Jacob Sten Petersen. Antigen-based Prediction and Prevention of Type 1 Diabetes. In Danish Medical Bulletin nr.4/2006 418-437.
6. Julio C. Delgado, Edmond J. Yunis. The Major Histocompatibility Complex and Disease. In Henry’s Clinical Diagnosis and Management by Laboratory Methods, Saunders-Elsevier, 21st Edition, 8, 894-902.
7. Laborator Synevo. Referintele specifice tehnologiei de lucru utilizate 2010. Ref Type: Catalog.
8. Robert J. Winchester. The Major Histocompatibility Complex. In Clinical Immunology. Principles and Practice, Mosby, Elsevier, Third Edition, 2008, 79-89.
9. V. Radha, K.S.Vimaleswaran, R.Deepa, V. Mohan. The genetics of diabetes mellitus. In Indian J Med Res 117/2003 225-238.
